3-1丂32嵶朎婜傑偱偺棏妱夁掱偺娤嶡偲奺妱媴偺柦柤
儚僇儗僆僞儅儃儎偺敪惗偺尋媶傪峴偆偵偁偨偭偰丄弶婜棏妱夁掱偺忣曬偼廳梫偱偡丅偟偐偟丄儚僇儗僆僞儅儃儎偺弶婜敪惗偵偮偄偰偼丄1910擭偺Delsman偺榑暥埲棃曬崘偝傟偰偄傑偣傫丅
Delsman偼丄僾儔儞僋僩儞僱僢僩傪梡偄偰奀偱嵦廤偟偰摼傜傟偨悢昐偺泱偺娤嶡傪尦偵偟偰偙偺榑暥傪敪昞偟傑偟偨丅偙偺拞偱斵偼丄儚僇儗僆僞儅儃儎偺尨挵娮擖廔椆傑偱偺徻嵶側棏妱僷僞乕儞偲妱媴攝抲偺婰嵹丄偦偟偰梒惗偵偄偨傞傑偱偺懱偺峔憿偵偮偄偰旕忢偵徻偟偔婰嵹傪偟偰偄傑偡丅偙傟偼丄摉帪偺尠旝嬀媄弍傪峫偊傞偲嬃偔傋偒傕偺偱偡丅傑偨丄斵偼丄儂儎偺曽朄偲偼堎側傞娐宍摦暔丒擃懱摦暔偺曽朄偵曧偭偰儚僇儗僆僞儅儃儎偺妱媴柦柤朄傪掕媊偟傑偟偨丅

巹偨偪偺尋媶幒偱偼丄Delsman偺曬崘傪妋擣偡傞偨傔丄嬤戙揑側曽朄傪梡偄偰儚僇儗僆僞儅儃儎偺弶婜棏妱夁掱傪挷傋傞偙偲偵偟傑偟偨丅嬶懱揑偵偼丄僲儅儖僗僉乕尠旝嬀偵傛傞棏妱夁掱偺幨恀嶣塭丒僞僀儉儔僾僗價僨僆嶣塭丄phallodin偵傛傞嵶朎枌愼怓泱偺尠旝嬀娤嶡丄傑偨弶婜泱偺棫懱夋憸峔抸偲偄偭偨曽朄偵傛傝丄弶婜棏妱夁掱偲尨挵娮擖偵偮偄偰徻偟偔尒偰峴偒傑偟偨丅
(1)僲儅儖僗僉乕尠旝嬀偵傛傞弶婜棏妱夁掱偺幨恀嶣塭
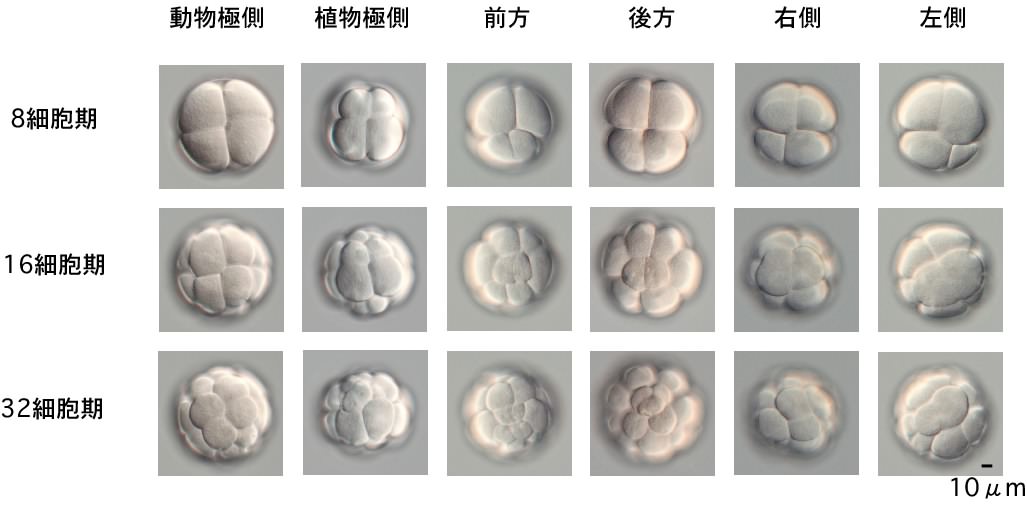
弶婜泱傪摦暔嬌懁丄怉暔嬌懁丄慜曽丄屻曽丄塃懁丄嵍懁偺慡俇曽岦偐傜弶婜棏妱夁掱偱妱媴攝抲偑偳偺傛偆偵曄壔偟偰峴偔偐傪丄僲儅儖僗僉乕尠旝嬀偵傛傝捛愓偟偰偄偒傑偟偨丅
(2)僲儅儖僗僉乕尠旝嬀偵傛傞僞僀儉儔僾僗價僨僆嶣塭
懕偄偰丄弶婜棏妱夁掱偺棏妱僷僞乕儞傪丄僲儅儖僗僉乕尠旝嬀偵傛傝僞僀儉儔僾僗價僨僆嶣塭偵傛傝妋擣偟傑偟偨丅偙偙偱偼丄弶婜棏妱偺條巕偲尨挵娮擖偺條巕傪嶣塭偟偨傕偺傪徯夘偟傑偡丅(價僨僆墈棗偵偼Quicktime Player偑昁梫偱偡)
儚僇儗僆僞儅儃儎偺尨挵娮擖偼旕忢偵憗偔丄32嵶朎婜偱巒傑傝傑偡乮儂儎偼110嵶朎婜乯丅
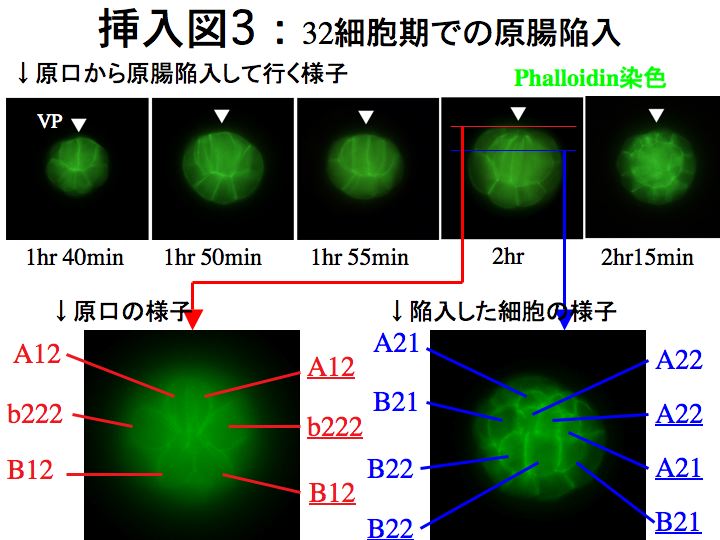
(3)嵶朎枌愼怓泱偺尠旝嬀娤嶡
偙偙偱偼丄弶婜泱偺嵶朎枌傪phalloidin偱愼怓偟丄偳偺傛偆偵妱媴偑尨挵娮擖偟偰偄偔偐傪娤嶡偟傑偟偨丅
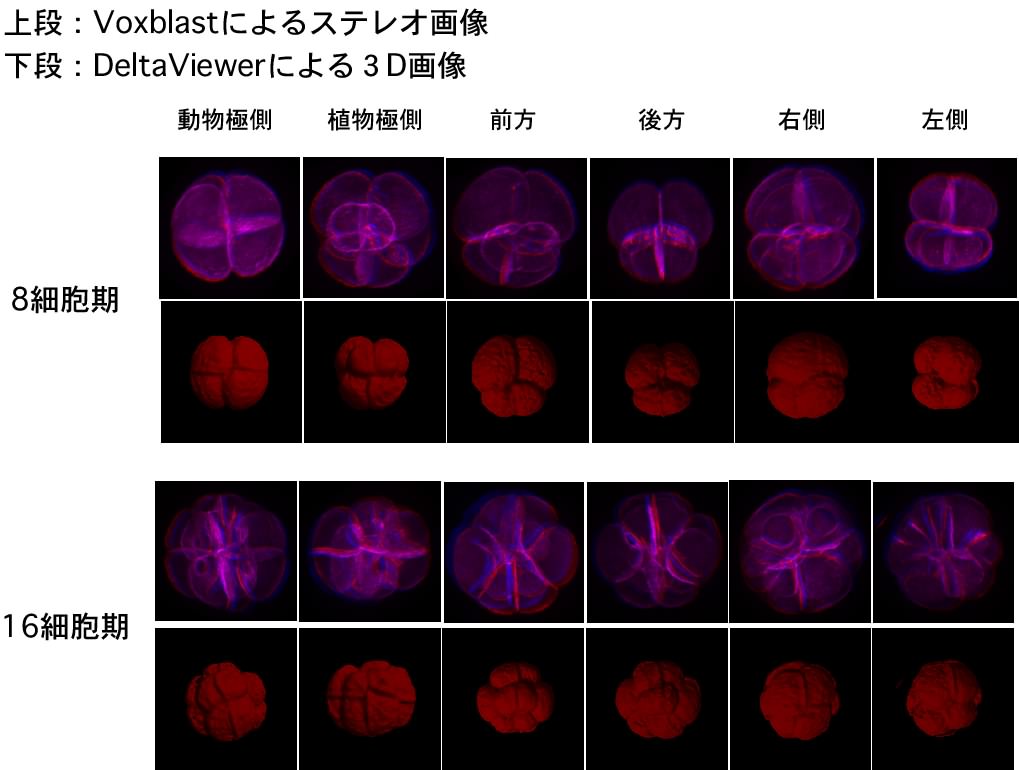
(4)嵶朎枌愼怓泱偺嫟徟揰尠旝嬀娤嶡偲棫懱夋憸峔抸
(3)偲摨條丄弶婜泱偺嵶朎枌傪phalloidin偱愼怓偟偨傕偺傪丄嫟徟揰尠旝嬀偱娤嶡屻丄夋憸張棟僜僼僩乮Voxblast丄DeltaViewer乯傪梡偄偰棫懱夋憸傪峔抸偟傑偟偨丅
偙偺傛偆偵丄32嵶朎婜傑偱偺棏妱僷僞乕儞偲妱媴攝抲偺妋擣偟丄奺妱媴偺柦柤乮柦柤偺巇曽偼Delsman偺曽朄偵曧偭偰偄傑偡乯傪峴偄傑偟偨丅偦偺寢壥丄Delsman偺曬崘偑傎傏惓偟偄偙偲偑暘偐傝傑偟偨丅
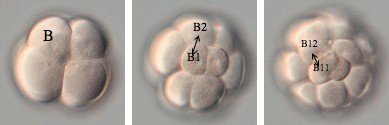
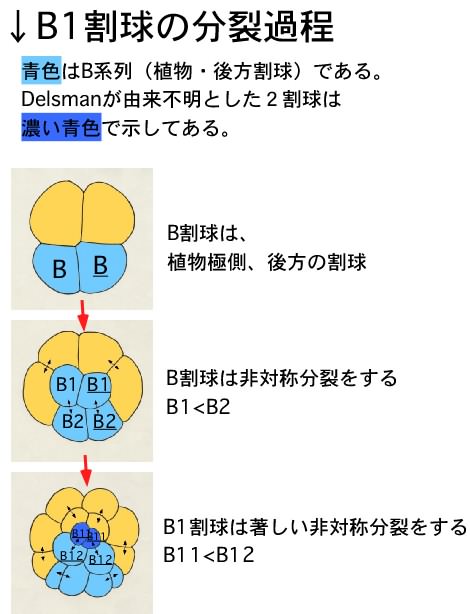
丒B1妱媴偺晄摍暘楐
偝偰丄Delsman偺曬崘偲堘偭偰偄偨偺偑丄B1妱媴偑挊偟偄晄摍暘楐傪峴偆偲偄偆揰偱偡丅Delsman偵傛傞偲丄B1妱媴偼暘楐偣偢丄偦偺屻30嵶朎婜偱桼棃晄徻偺2妱媴偑尰傟傞偲彂偐傟偰偄傑偡丅偟偐偟変乆偑娤嶡偟偨偲偙傠丄B1偑挊偟偄晄摍暘楐傪偟偨寢壥偑丄偙偺2妱媴偱偁傞偲暘偐傝傑偟偨丅B1妱媴偼儚僇儗僆僞儅儃儎泱偺嵟屻曽妱媴偱偁傝丄偙偺傛偆偵嵟屻曽妱媴偑挊偟偄晄摍暘楐傪偡傞偲偄偆揰偼丄儅儃儎偲帡偰偄傑偡丅
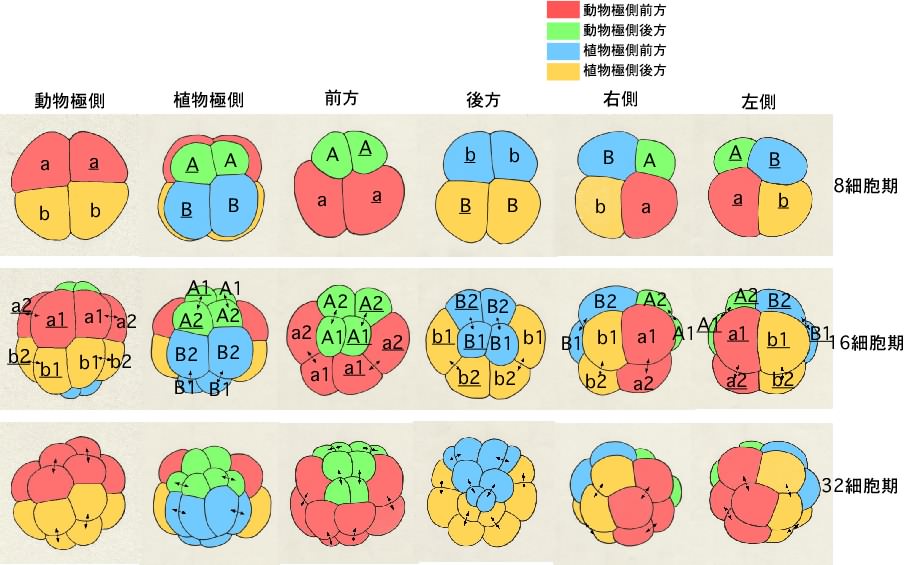
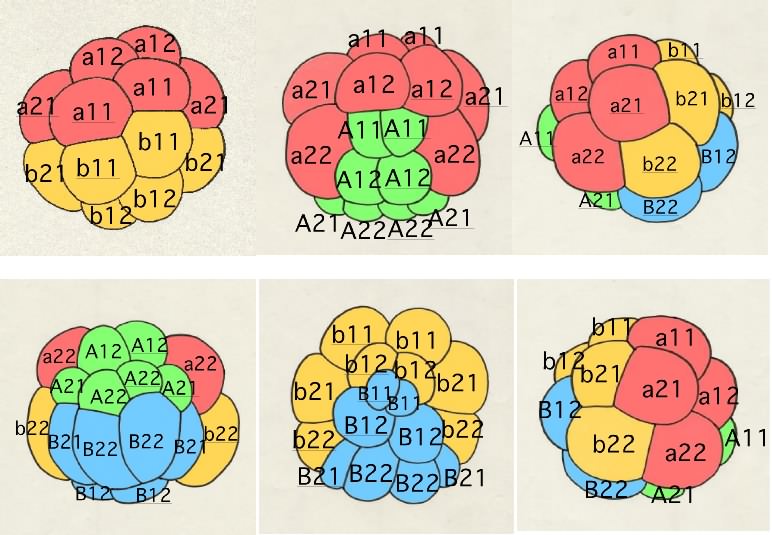
仏儚僇儗僆僞儅儃儎偺妱媴柦柤朄
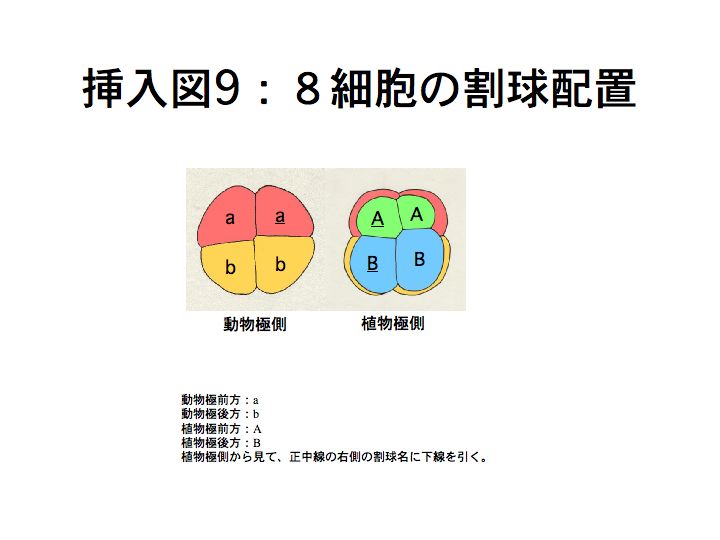
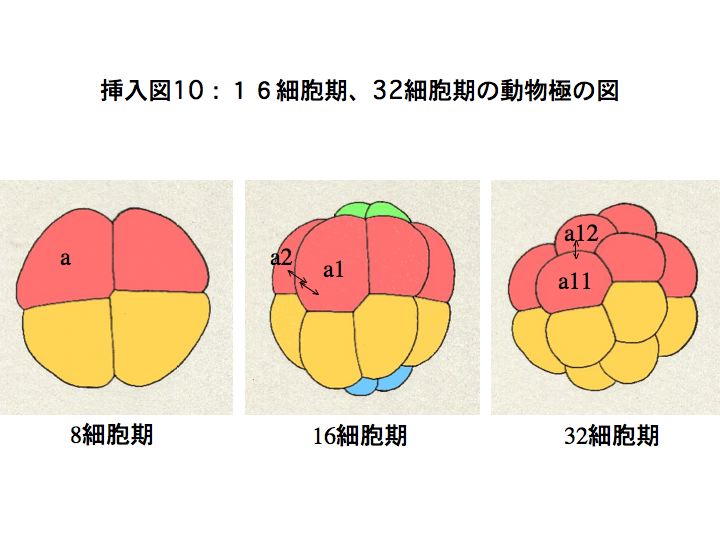
奺妱媴偼俉嵶朎婜偐傜幆暿偱偒傞傛偆偵側傝丄妱媴柤偑晅偒傑偡丅
俉嵶朎婜偱偼丄偦傟傑偱嬒摍偱偁偭偨係偮偺妱媴偵戝偒偝偺堘偄偑惗偠偼偠傔傑偡丅摦暔嬌懁偼戝偒偔偰丄傎傏嬒摍側係妱媴偐傜側傝丄怉暔嬌懁偼摦暔嬌懁傛傝彫偝偄係妱媴偐傜側傝傑偡丅偝傜偵丄怉暔嬌懁偱偼丄慜曽偺俀妱媴偺曽偑彫偝偔側傝傑偡丅摦暔嬌懁慜曽妱媴傪a丄屻曽妱媴傪b丄怉暔嬌懁慜曽妱媴傪A丄屻曽妱媴傪B偲屇傃傑偡丅傑偨丄惓拞慄傪嫴傫偱塃懁偺妱媴偵壓慄傪堷偒傑偡丅
俉嵶朎婜偐傜侾俇嵶朎婜偵偐偗偰偺暘楐偱偼丄椺偊偽丄摦暔嬌懁慜曽妱媴偺a妱媴偼乭a1乭偲乭a2乭偵暘楐偟傑偡丅暘楐屻偺柡嵶朎偺偆偪丄摦暔嬌偵嬤偄曽偵1丄怉暔嬌偵嬤偄曽偵2傪偮偗傑偡丅偝傜偵丄侾俇嵶朎婜偐傜32嵶朎婜偵偐偗偰偺暘楐偱偼丄a1妱媴偼摦暔嬌偵嬤偄曽偺乭a11乭偲怉暔嬌偵嬤偄曽偺乭a12乭偵暘楐偟傑偡丅
嶲峫暥專
- H. C. Delsman丄BEITRÄZGE ZUR ENTWICKLUNGSGESCHICHITE VON OIKOPLEURA DIOICA丄Verch. Rijksinst. Onderz. Zee, 3 (2), 1-24 (1910)