

研究内容
2.光合成反応中心にリンクする電子伝達経路
2-1. はじめに
キノール:シトクロムc酸化還元酵素は、光合成系のみならずミトコンドリアの呼吸鎖電子伝達系にも存在しています。キノールの酸化に伴うプロトン濃度勾配の形成が、ATP 合成酵素の駆動力となっています。キノール:シトクロムc酸化還元酵素はそのサブユニット構成から、紅色光合成細菌ではチトクロム b/c1 複合体(Rieke Fe-S タンパク、チトクロム b、チトクロム c1)、ラン色細菌や高等植物のチラコイド膜ではチトクロム b6/f 複合体(Rieke Fe-S タンパク、チトクロム b6、チトクロム f、サブユニットIV)と呼ばれています。しかし残念ながら緑色イオウ細菌やヘリオバクテリアのキノール:シトクロムc酸化還元酵素は未だに単離されていません。おそらく膜タンパク質複合体としてきわめて不安定であるためなんだと思います。これについては将来の課題として残されていて、私たちはいずれこの課題に取り組んでいきたいと日々アイデアをあたためています。
膜結合型チトクロムc
これまでの研究の中で、膜タンパク質間の電子伝達に機能する膜結合型チトクロムcの重要性に気づいて研究を大きく展開させています。緑色イオウ細菌やヘリオバクテリアの反応中心複合体とキノール酸化還元酵素は膜結合型チトクロムcにより直接つながっていて、スーパーコンプレックス(超分子複合体)を形成しているものと想像されます。このような電子伝達共役系を明らかにすることは、光合成電子伝達系の進化的成立過程をより深く考察していく上で、非常に重要かつ興味深い問題であると考えています。
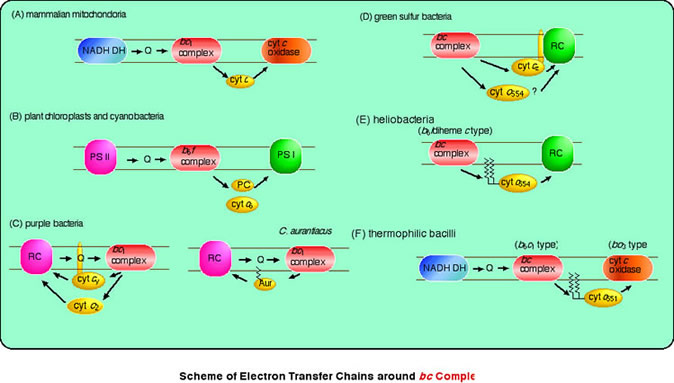
2-2. 緑色イオウ細菌の電子伝達経路
光合成反応中心が光エネルギーを吸収し、効率よく還元力を作り出すためには、常に反応中心がturn overする反応機構が重要です。つまり光によって酸化された反応中心(酸化側)には速やかに電子の供給が行われ、再還元されることが必要です。この経路は高等植物の葉緑体やシアノバクテリアではかなり調べられていて、キノール:プラストシアニン酸化還元酵素(シトクロムb6/f複合体)と光化学系I反応中心の間には、水溶性のプラストシアニン(PC)やシトクロムc6(cyt c6、シアノバクテリアのみ)が働いています。今では、PCやcyt c6、およびシトクロムb6/f複合体、系I反応中心複合体の詳細な立体構造も分かっていますので、分子レベルでの反応機構を議論することも可能となっています。しかしながら緑色イオウ細菌やヘリオバクテリアの反応中心に電子を渡す経路については、実はあまりよく分かっていません。紅色細菌や植物(シアノバクテリア)のもつ電子伝達経路との比較の上で推測されているに過ぎません。特に緑色イオウ細菌が還元型イオウ(S-, S2O32-, S)を硫酸イオン(SO42-)にまで酸化する経路は未だに不明であり、このイオウ代謝経路は地球上のイオウ循環経路を考える上で非常に重要な位置を占めています。最近ではさまざまなイオウ細菌のゲノム解析からイオウ酸化機構が議論されるようになりましたので、将来の発展が楽しみな領域です。私たちも少しずつではありますが、化学反応としても非常に興味深いイオウ酸化系を研究していきたいと考えています。
再還元経路に関する研究
すでに我々は反応中心の再還元経路として、膜結合型シトクロムcz(cyt cz)が大きく揺らぎながら直接シトクロムbc複合体から電子を受け取っていることを証明している(1998年 Biochemistry)。今回は1970年代から研究されている低分子量の水溶性シトクロムc-554(cyt c-554)の機能について、試験管内再構成実験系を用い、閃光照射による過渡吸収測定により詳細に検討した。その結果、これまで推測されていたように、cytc-554がシトクロムbc複合体と反応中心との間の電子伝達体として機能しているのではなく、まったく独立的に反応中心cyt czに電子を供与しているということが判明した。cyt c-554は、おそらくチオ硫酸酸化系(Sox系)から直接電子を受容し、光合成反応系に電子を運ぶ伝達体として機能しているのであろう。これにより、早晩、微生物学に長年にわたって記載されてきた緑色イオウ細菌の光合成電子伝達経路が書きかえられるものと思われる。ある意味、古くて新しい話題であり、光合成細菌研究の歴史の上では重要な発見であると考えている。
(参考文献)Y. Tsukatani, C. Azai, T. Kondo, S. Itoh and H. Oh-oka (2008) Parallel electron donation pathways to cytochrome cz in the type I homodimeric photosynthetic reaction center complex of Chlorobium tepidum Biochim. Biophys. Acta 1777: 1211-1217.

(説明)閃光照射後のcyt bの酸化キネティックス:cyt c-554/555存在下では、antimycin Aを加えるとcyt bの酸化量は半減する。このことは、cyt c-554/555がシトクロムbc複合体から電子を受け取らないことを示している。
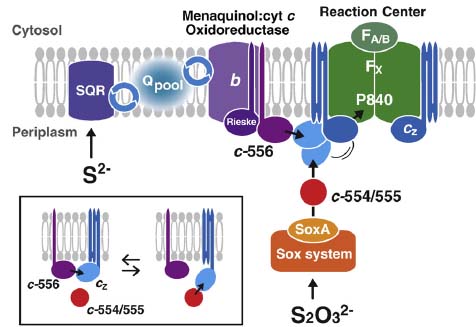
(説明)緑色イオウ細菌の反応中心の再還元側では、2つの独立した電子供与経路が存在することを示している。
cyt c-554の機能について再考察
先に我々はcyt c-554がチオ硫酸酸化系(Sox系)から直接電子を受容し、光合成反応系に電子を運ぶ伝達体として機能していると報告した(BBA, 2008:下記参照)。この結論自体は決して間違いではないが、意外なことにcyt
c-554欠損株においてもチオ硫酸を完全酸化する能力を持ち、野生株と同程度に生長することを見出した。このことはチオ硫酸酸化系(Sox系)と光合成反応中心との間には別の電子伝達経路(あるいは未知の電子伝達成分)が存在していることを示す重要な発見であり、緑色イオウ細菌における低分子量水溶性シトクロムc(cyt c-554/5555)の機能について再考察する必要性がある。チオ硫酸を酸化することができないいChlorobium ferroxidansは還元鉄(Fe2+)を電子源として生育するが、ゲノム上にはcyt c-554/555をコードしていることが分かっている。cyt c-554/555の関与する電子伝達経路は、1970年代にはすでに明らかとなったと思われていた常識である。覆された常識は、果たしてこの先、どのような結末を迎えるのであろうか?
(参考文献)C. Azai, Y. Tsukatani, J. Harada and H. Oh-oka (2009) Sulfur oxidation in mutants of the photosynthetic green sulfur bacterium Chlorobium tepidum devoid of cytochrome c-554 and SoxB Photosynth. Res. 100: 57-65.
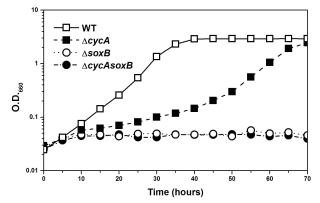
(説明)cyt c-554欠損株(ΔcycA)は野生株(WT)と比較すると生長速度は緩やかであるがチオ硫酸を完全酸化することができ、野生株とほぼ同程度まで増殖する。チオ硫酸酸化に関与するSoxB欠損株(ΔsoxB)はチオ硫酸を酸化できず、増殖しない。