染色体の安定維持機構
ヒトをはじめ多くの生物種のゲノム配列は解読されました。たった1つの受精卵が分裂を繰り返し、約60兆個の細胞になって人体を形作ります。それら1つ1つの細胞が同じ遺伝情報を持つことから、遺伝情報を維持するための驚異的なメカニズムが存在することが分かります。しかし、遺伝情報を担う染色体がどのようにして正確に複製、維持されて、均等に娘細胞に受け継がれているのかは今も解明されていません。そこで、我々は染色体の安定維持機構を明らかにするために分裂酵母やヒト培養細胞を用いて、DNA複製、組換え、修復、転写、エピジェネティック制御、染色体構造、染色体分配などについて研究を行っています。
セントロメアに於ける相同組換えの役割
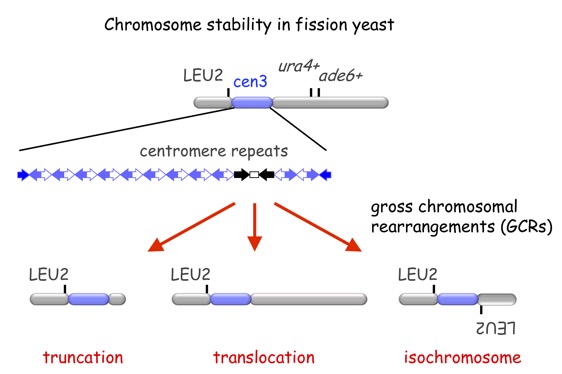
染色体の安定維持機構を研究するために、我々は、異常染色体の発生頻度を測定し、その構造を明らかにすることが出来る実験系を分裂酵母を用いて開発しました(Nakamura et al. 2008)。この系を用いた解析により、細胞周期チェックポイント制御やDNA相同組換えが染色体異常の抑制に重要な役割を果たすことを明らかにしました。また、興味深いことに、相同組換え因子Rad51を遺伝子破壊すると分裂酵母のセントロメア領域を構成する逆向きの反復配列を「のりしろ」にして両方の染色体腕の配列が同一の同腕染色体が高頻度に形成されることが明らかになった。これらの結果から、Rad51が関与する相同組換えはセントロメア・リピート配列を介した染色体異常を抑制すると考えられる。染色体のセントロメア領域は分裂期微小管が結合する動原体が形成されまた、CENP-Aクロマチンやヘテロクロマチンなどの特徴的なクロマチン構造が形成れます。また、ヒトを含む多くの真核生物のセントロメアDNAはリピート配列により構成されている。分裂(M)期微小管と染色体との接着部位としてセントロメアは染色体分配をつかさどることから、ゲノム維持に中心的な役割を果たす染色体領域であると言えます。ところが、セントロメアは染色体異常が起こりやすい染色体脆弱部位でもあります。したがって、生物はセントロメア領域を維持するための特別なシステムを備えていると考えられます。我々は相同組換え因子Rad51が細胞周期の複製(S)期特異的にセントロメアに結合し、セントロメアの再編を抑制することを明らかにしました。このことから、相同組換えはセントロメア領域の維持で重要であると考えられます。分裂酵母のセントロメアは、比較的単純なリピート配列により構成されていることから様々な改変を施すことができます。また、高等真核生物と共通したクロマチン構造をつくります。このようにセントロメア研究に最適なモデル生物である分裂酵母を用いて、セントロメア組換えの分子機構とその生理的意義の解明を目指しています。
DNA複製の制御機構
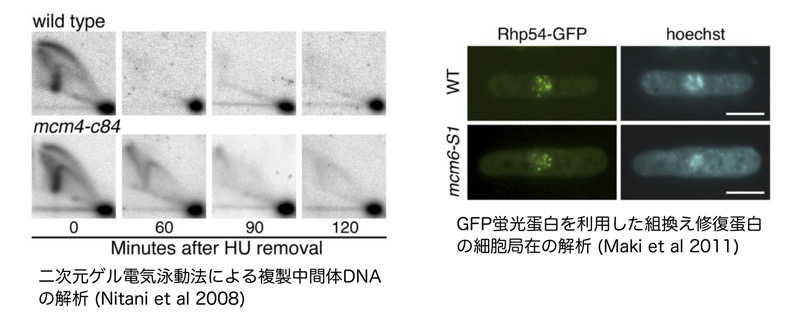
真核生物の染色体上には多数の複製起点が存在し、その数はゲノム全体で数百以上あります。ただし、個々の細胞を見たとき、それら全ての複製起点から複製開始(発火)する訳ではなく実際に発火が起こるのはその一部であり大部分の複製起点からの発火は起きこりません。この「複製起点パラドクス」と呼べる現象がなぜ起こるのか。その答えは未だ明らかではなませんが、複製起点の数が減少すると複製ストレス感受性の増大、染色体異常、発生異常、がん化など様々な問題が生じることから、生理的に重要な意義を担っていると考えられています。
遺伝情報を正確に娘細胞に伝えるためには、その担い手である染色体DNAを全長に渡って正確に複製することが必要です。このときDNA分子だけでなくヒストン修飾などのエピジェネティックなゲノム情報も正確にコピーする必要があります。また、様々な内的・外的要因により複製の進行は阻害される為、複製フォーク(複製中間体)はこれらの状況にも適切に対応して染色体の倍加を達成しなければならない。このように、DNA複製は実に複雑なことをやり遂げているが、その分子メカニズムは謎のままである。我々は複製起点の構成因子であり複製フォークの駆動部でもあるMcm2–7ヘリケースに着目して「DNA複製によるゲノム維持」の分子メカニズムを明らかにしたいと考えています。