
研究概要
松野研究室では、動物のからだが決まった形になる仕組みを、細胞や遺伝子の機能のレベルで理解したいと考え、次のような研究を行っています。形態形成と細胞シグナル~Notchシグナルを解明する~
胚や器官の形態が形成されるためには、それらにおいてパターンが形成されることが必要です。パターンのおおもとは、卵の中における遺伝子産物の不均一な分布です。パターンは、位置情報を担っており、それにしたがって、細胞の分化、増殖、細胞死、移動などが起こります。
パターンの形成は、ドミノ倒し式に連続して起こり、パターンが細分化されていきます。このとき、パターンの受け渡しや、受け取りに必要なのが、細胞シグナルです。
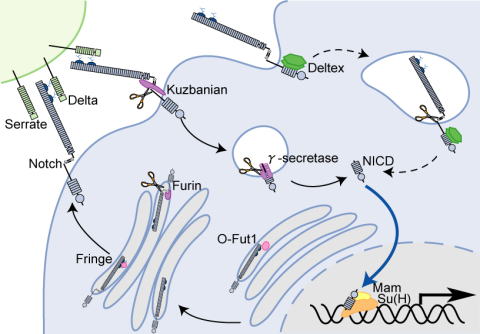
松野研究室では、細胞シグナルの中でも、Notchシグナルに焦点を絞って研究を進めてきました。Notchシグナルは、細胞間の直接的接触を介する細胞シグナルで、発生や恒常性の維持に重要な機能をはたしています。Notchシグナルは、ショウジョウバエからヒトまで進化的に保存されており、ヒトでは、ガンや各種遺伝病との関連が明らかにされています。松野は、当時エール大学(現ハーバード大学)にいたSpyros Artavanis-Tsakonas教授のもとに留学し、日本人としては初めて、Notchシグナルの研究に従事しました。それ以降も、松野研究室では、Notchシグナルの伝達に必要な新たな遺伝子の同定に成功しています。松野研究室では、今後も、Notchシグナルの研究を進めていきます。 Notchシグナルの異常は、ガン化と密接な関係にあります。ショウジョウバエを用いてNotchシグナルの伝達機構やその制御法を明らかにすることで、医学への応用が可能な成果が得られます。
松野研究室では、ショウジョウバエ遺伝学の長所を活かして、Notchシグナルの新規な構成遺伝子の探索を続けています。現在でも、機能既知なNotchシグナル構成遺伝子の同定に成功しており、解析を行っています。
細胞膜受容体は、糖鎖修飾を受けています。しかし、受容体の成熟や機能におけるそれぞれの糖鎖修飾の機能には未知な点が多く残されています。Notchの細胞ドメインに存在するepidermal growth factor様反復配列も糖鎖修飾を受けています。松野研究室では、この糖鎖修飾のうち、O型糖鎖の機能に興味をもって研究を進めています。
我々の最近の研究から、小胞体ストレス応答に関連すると考えられるプロセスが、Notchシグナルに必要であることが示唆されています。この機能は、Notchシグナルの構成因子である、複数回膜貫通タンパク質Pecanexの機能が欠失すると損なわれます。現在、pecanex遺伝子の機能に関する研究を進めています。
パターンの形成は、ドミノ倒し式に連続して起こり、パターンが細分化されていきます。このとき、パターンの受け渡しや、受け取りに必要なのが、細胞シグナルです。
松野研究室では、細胞シグナルの中でも、Notchシグナルに焦点を絞って研究を進めてきました。Notchシグナルは、細胞間の直接的接触を介する細胞シグナルで、発生や恒常性の維持に重要な機能をはたしています。Notchシグナルは、ショウジョウバエからヒトまで進化的に保存されており、ヒトでは、ガンや各種遺伝病との関連が明らかにされています。松野は、当時エール大学(現ハーバード大学)にいたSpyros Artavanis-Tsakonas教授のもとに留学し、日本人としては初めて、Notchシグナルの研究に従事しました。それ以降も、松野研究室では、Notchシグナルの伝達に必要な新たな遺伝子の同定に成功しています。松野研究室では、今後も、Notchシグナルの研究を進めていきます。 Notchシグナルの異常は、ガン化と密接な関係にあります。ショウジョウバエを用いてNotchシグナルの伝達機構やその制御法を明らかにすることで、医学への応用が可能な成果が得られます。
松野研究室では、ショウジョウバエ遺伝学の長所を活かして、Notchシグナルの新規な構成遺伝子の探索を続けています。現在でも、機能既知なNotchシグナル構成遺伝子の同定に成功しており、解析を行っています。
細胞膜受容体は、糖鎖修飾を受けています。しかし、受容体の成熟や機能におけるそれぞれの糖鎖修飾の機能には未知な点が多く残されています。Notchの細胞ドメインに存在するepidermal growth factor様反復配列も糖鎖修飾を受けています。松野研究室では、この糖鎖修飾のうち、O型糖鎖の機能に興味をもって研究を進めています。
我々の最近の研究から、小胞体ストレス応答に関連すると考えられるプロセスが、Notchシグナルに必要であることが示唆されています。この機能は、Notchシグナルの構成因子である、複数回膜貫通タンパク質Pecanexの機能が欠失すると損なわれます。現在、pecanex遺伝子の機能に関する研究を進めています。
左右極性から左右非対称な形態形成へ~細胞生物学と力学の融合~

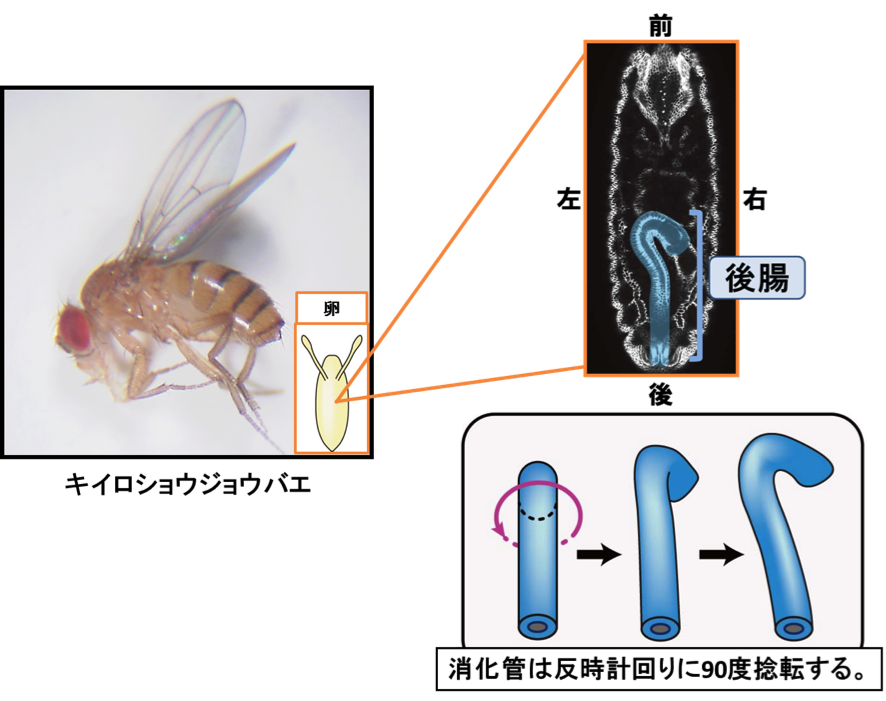
動物のからだが左右非対称なことがよくあります。例えば、左に眼があるのがヒラメ、右に眼があるのが カレイです。また、ヒトの内臓器官のほとんどは左右非対称で、ご存知のように、心臓は左側にあります。からだが左右非対称になる仕組みについては長い間の謎でしたが、日本のグループの活躍によって、脊椎動物の一部ではその仕組みがよく理解されるようになりました。しかし、左右非対称性が形成される仕組みは動物のグループで異なり、無脊椎動物では脊椎動物とは異なる仕組みで左右非対称性がつくられていると考えられています。
松野研究室では、無脊椎動物で働いている、左右非対称性形成のこれまでに知られていない仕組みを明らかにしたいと考え、分子遺伝学の研究に適したショウジョウバエを用いて研究を行っています。ショウジョウバエでは、消化管、精巣、脳、雄の生殖器に左右非対称性が認められます。野生型では、これらの左右非対称性が乱れることはほとんどなく、左右非対称性が遺伝的に決められていることがわかります。我々は、発生過程で最初に左右非対称性を示す、胚消化管に注目して研究を行ってきました。胚消化管の後部にある後腸は、左右非対称な形態をとる過程で左ネジ方向に90度捻転します。遺伝学を用いたこれまでの研究で、後腸の左右非対称性の形成に必要な遺伝子の同定に成功しており、I型ミオシンやアクチン細胞骨格に依然した、これまでに知られていない左右非対称性な形態形成の機構を明らかにしました。
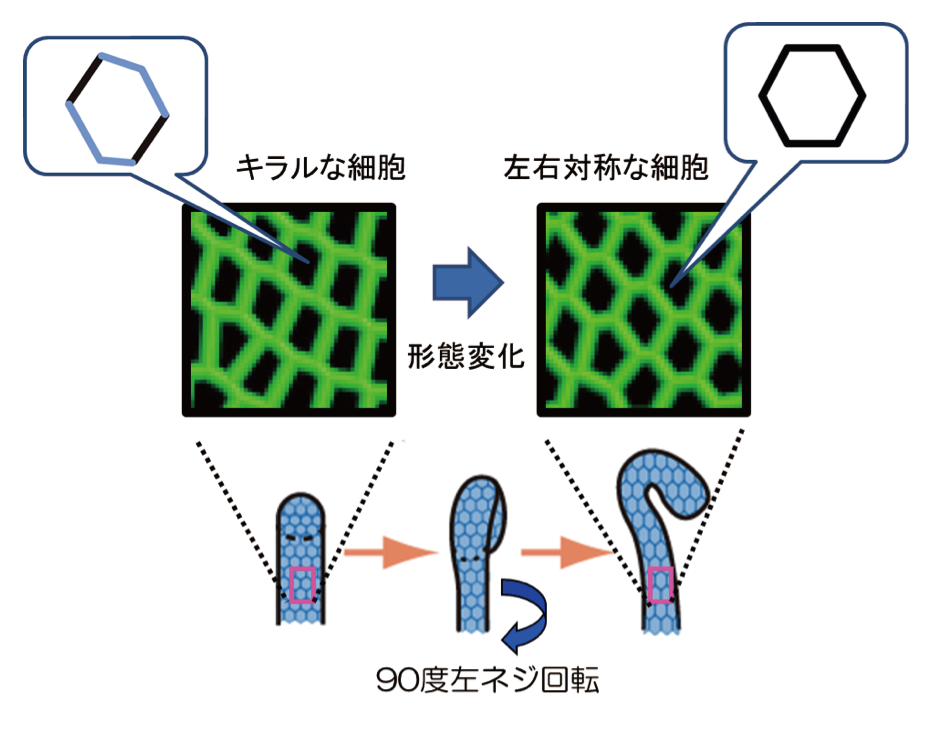
特に重要な発見は、この捻転が起こる前に、消化管の内壁をつくる上皮細胞の管内面側の形が左右に歪むことを明らかにしたことです。この左右に歪んだ形は、鏡に映した像が元の像と重ならないことから、“キラル”な性質をもっています。我々は、これを平面内細胞キラリティと呼ぶことにしました。平面内細胞キラリティは、松野研究室が世界で初めて明らかにしたものです。平面内細胞キラリティは、消化管の捻転に伴って解消されました。平面内細胞キラリティのコンピュタ・シミュレーションを行うと、平面内細胞キラリティによって消化管の左ネジ90度捻転が説明できることがわかりました。これらの研究から、組織の形がつくられる新たな仕組みを明らかにできました。別のグループの研究から、平面内細胞キラリティが哺乳類の培養細胞でもみつかりました。つまり、ショウジョウバエを用いた我々の研究によって、動物に普遍的な新しい細胞の極性が見つけられました。
平面内細胞キラリティによって後腸の左ネジ90度捻転が説明できることから、松野研究室では、これまであまり注目されてこなかった、細胞が生み出す「力」による組織の変形に焦点をあて、組織形態の左右非対称な変化を理解したいと考えています。このような「力」を生み出している遺伝子の働きを調べるためには、「力」を定量的に扱うことが必要です。そこで、この捻転の際に発生する回転トルクを測定することにしました。磁気ビーズをショウジョウバエ後腸に顕微注入し、磁気ピンセットで磁力を加え、捻転の回転を人為的に止める方法を用いて、捻転トルクの測定に成功しています。トルクを定量的に扱えるようになったことから、今後は、「力」の発生や、「力」の方向の決定に必要な遺伝子の研究が可能になると考えています。つまり、今後の研究では、これまでに行われたことのない、「力」の遺伝学的解析を行っていきます。
後腸の捻転は、常に90度です。我々は、「力」の効果が何らかの方法で読み取られ、捻転の停止を誘発している可能があると考えています。つまり、組織が左右極性などのパターンにもとづいて「力」を発生し、その「力」で組織・器官の形態は変化し、これによって次のパターン形成などの応答が起こっていると予測しているのです。そこで、細胞・組織が「力」を発生する機構や、その「力」が細胞に感知される機構を調べていきます。我々は、「力」が、発生プログラムの要素であると捉えています。松野研究室では、これを今後の重要なテーマとして位置付けています。
松野研究室では、無脊椎動物で働いている、左右非対称性形成のこれまでに知られていない仕組みを明らかにしたいと考え、分子遺伝学の研究に適したショウジョウバエを用いて研究を行っています。ショウジョウバエでは、消化管、精巣、脳、雄の生殖器に左右非対称性が認められます。野生型では、これらの左右非対称性が乱れることはほとんどなく、左右非対称性が遺伝的に決められていることがわかります。我々は、発生過程で最初に左右非対称性を示す、胚消化管に注目して研究を行ってきました。胚消化管の後部にある後腸は、左右非対称な形態をとる過程で左ネジ方向に90度捻転します。遺伝学を用いたこれまでの研究で、後腸の左右非対称性の形成に必要な遺伝子の同定に成功しており、I型ミオシンやアクチン細胞骨格に依然した、これまでに知られていない左右非対称性な形態形成の機構を明らかにしました。
特に重要な発見は、この捻転が起こる前に、消化管の内壁をつくる上皮細胞の管内面側の形が左右に歪むことを明らかにしたことです。この左右に歪んだ形は、鏡に映した像が元の像と重ならないことから、“キラル”な性質をもっています。我々は、これを平面内細胞キラリティと呼ぶことにしました。平面内細胞キラリティは、松野研究室が世界で初めて明らかにしたものです。平面内細胞キラリティは、消化管の捻転に伴って解消されました。平面内細胞キラリティのコンピュタ・シミュレーションを行うと、平面内細胞キラリティによって消化管の左ネジ90度捻転が説明できることがわかりました。これらの研究から、組織の形がつくられる新たな仕組みを明らかにできました。別のグループの研究から、平面内細胞キラリティが哺乳類の培養細胞でもみつかりました。つまり、ショウジョウバエを用いた我々の研究によって、動物に普遍的な新しい細胞の極性が見つけられました。
平面内細胞キラリティによって後腸の左ネジ90度捻転が説明できることから、松野研究室では、これまであまり注目されてこなかった、細胞が生み出す「力」による組織の変形に焦点をあて、組織形態の左右非対称な変化を理解したいと考えています。このような「力」を生み出している遺伝子の働きを調べるためには、「力」を定量的に扱うことが必要です。そこで、この捻転の際に発生する回転トルクを測定することにしました。磁気ビーズをショウジョウバエ後腸に顕微注入し、磁気ピンセットで磁力を加え、捻転の回転を人為的に止める方法を用いて、捻転トルクの測定に成功しています。トルクを定量的に扱えるようになったことから、今後は、「力」の発生や、「力」の方向の決定に必要な遺伝子の研究が可能になると考えています。つまり、今後の研究では、これまでに行われたことのない、「力」の遺伝学的解析を行っていきます。
後腸の捻転は、常に90度です。我々は、「力」の効果が何らかの方法で読み取られ、捻転の停止を誘発している可能があると考えています。つまり、組織が左右極性などのパターンにもとづいて「力」を発生し、その「力」で組織・器官の形態は変化し、これによって次のパターン形成などの応答が起こっていると予測しているのです。そこで、細胞・組織が「力」を発生する機構や、その「力」が細胞に感知される機構を調べていきます。我々は、「力」が、発生プログラムの要素であると捉えています。松野研究室では、これを今後の重要なテーマとして位置付けています。