1 オタマボヤの紹介
私たちの研究室では、ワカレオタマボヤ(Oikopleura dioica)という生物を研究の対象としています。ここでは、オタマボヤがどういう生物なのかを紹介します。
近年、ワカレオタマボヤ(Oikopleura dioica)の継代飼育が研究室内でできるようになり、オタマボヤを用いた研究への可能性は大きく広がりました。 ワカレオタマボヤは実験動物として以下のような特徴をもち、発生生物学において新たなモデル生物となる可能性を備えていると考えられます。
- 研究室での継代飼育が可能
- ライフサイクルが非常に短く、18℃で飼育した場合5日で卵や精子を産む
- 世界中でみられる種であり、日本での採集も容易である
- 卵や胚が透明で顕微鏡観察に適している
- 胚細胞の数が少ない
- 成体の構造が単純である
- 脊索動物に共通の特徴を終生保持し、他の脊索動物との比較が可能である
- ゲノムプロジェクト、ESTプロジェクトが進行している
- ゲノムがコンパクトで遺伝子間距離が短い(5kbに1遺伝子)
- 遺伝子重複がほとんどない
1-1 オタマボヤのかたち
下図はオタマボヤの半模式図です。オタマボヤの体はオタマジャクシのような形をしていて、大きく体幹部と尾部に分けられます。体幹部には、口や腸、心臓、生殖巣など、尾部には脊索や筋肉などが含まれています。体の全体は、ハウスと呼ばれる構造物で覆われています。
オタマボヤの細胞数は非常に少なく、例えばワカレオタマボヤの筋肉細胞は尾部に20個、脊索細胞は20個です。
オタマボヤは他の尾索動物と異なり変態後も尾を持ち続けます。このため、脊索動物に共通の特徴である脊索を終生保持し続けます。
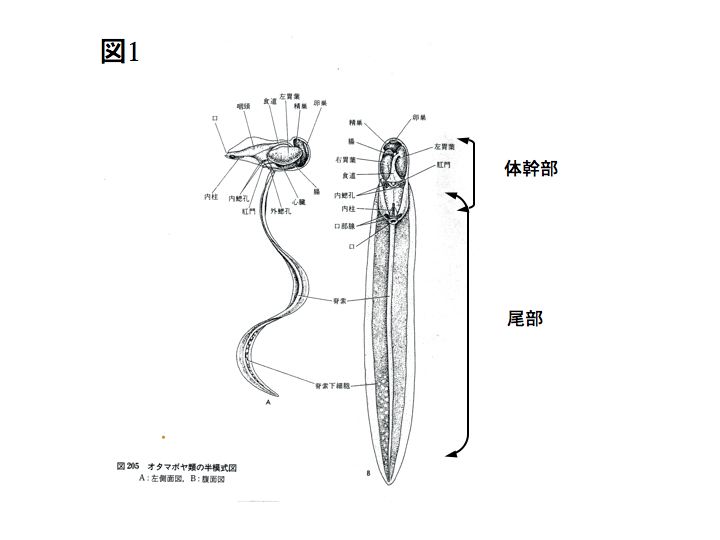
(動物系統分類学8(下)中山書店より)
1-2 生態について
オタマボヤは世界中の海に生息する重要な動物プランクトンとして、生態学的研究がなされています(Hirst et al.,2003)。バイオマスが海洋で2番目に多いグループで、マリンスノウの主となる構成成分であることが知られています。主に、海水の上層(〜100m)に多く生息していますが、600〜3600mの深さにもいることが確認されています。ほとんどのオタマボヤの体の大きさは数ミリメートルですが、中には体長が8〜9cmでハウスが 1mになるもの(Bathochordaeinae)もいます。
海の中では、ヒラメ幼魚がワカレオタマボヤを摂食していることがShelbourne(1962)により報告されています。オタマボヤの生態学的重要性のひとつは、魚が餌として利用できない小さな生物をオタマボヤが食べ、オタマボヤを魚が食べることによって食物連鎖のひとつの鎖となることです。また、捨てられたハウスは甲殻類のコペポーダによって食されていることが報告されています(Alldredge,1972)。フロリダの10〜15mの深さでの観測によると、1m3あたり平均329.8個のハウスが見つかっており、オタマボヤ類による赤潮の発生がいくつか記録されています(Seki,1973;Alldredge,1982)。
私たちの研究対象であるワカレオタマボヤは広く世界の海域、特に沿岸内域に密集しており、広温・広塩性の内湾種だと考えられます。ワカレオタマボヤの大きさは体幹部0.5〜1.3mm、尾部2.0〜3.9mmです。日本の沿岸にも多くみられ、私たちはこれまで、神奈川県小網代沖、和歌山県田辺湾沖、大阪府大阪湾で採集しています。
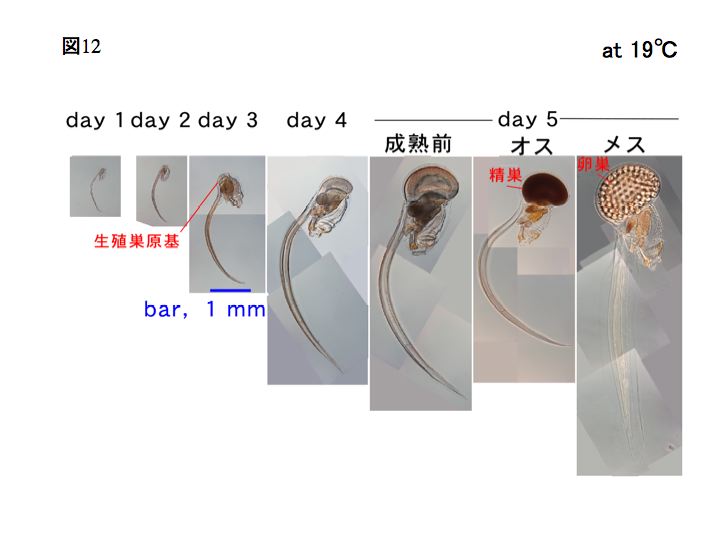
オタマボヤの一生はとても短く、ワカレオタマボヤの場合ではおおよそ、15℃で7日間、19℃で5日間、25℃では3日間です。19℃の場合、受精後から幼生になって孵化まで3時間、変態するまで10時間です。
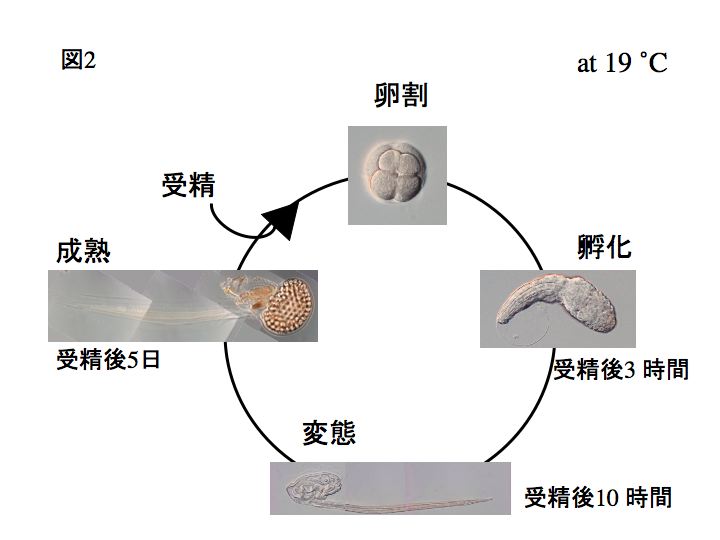
また、ワカレオタマボヤはオタマボヤ類の中で唯一、雌雄異体の種です(ワカレオタマボヤ以外の尾索動物は知られている限り雌雄同体です)。左下の写真は雌、右下は雄です。それぞれ、卵巣と精巣が発達しています。
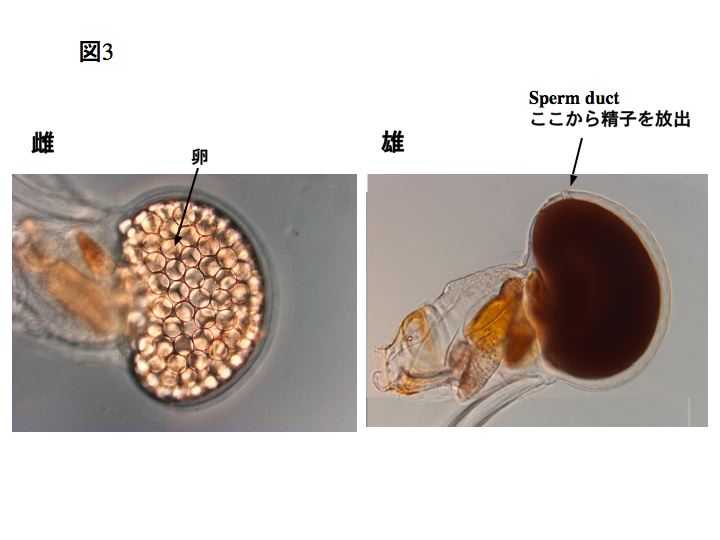
1-3 分類について
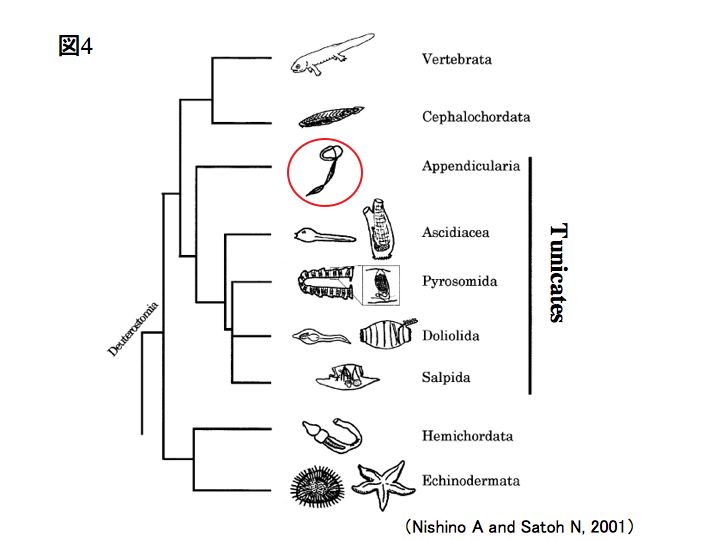
オタマボヤは、私たちヒトと同じ脊索動物門に属しており、尾索動物亜門オタマボヤ綱に分類される生物です。オタマボヤ綱には3つの科(オタマボヤ科・サイズチボヤ科・カサオタマボヤ科)があり、ワカレオタマボヤはオタマボヤ科に属しています。
- Chordata(脊索動物門)
- Vertebrata(脊椎動物亜門)
- Cephalochordata(頭索動物亜門)
- Urochordata(尾索動物亜門)
- Ascidiacea(ホヤ綱)
- Thaliacea(タリア綱)
- Appendicularia(オタマボヤ綱)
- Oikopleuridae(オタマボヤ科)...Oikopleura dioica(ワカレオタマボヤ)
- Fritillaridae(サイズチボヤ科)
- Kowalevskiidae(カサオタマボヤ科)
(岩波生物学辞典 第四版 岩波書店;動物系統分類学8(下)中山書店より)
1-4 オタマボヤ類の体の構造について
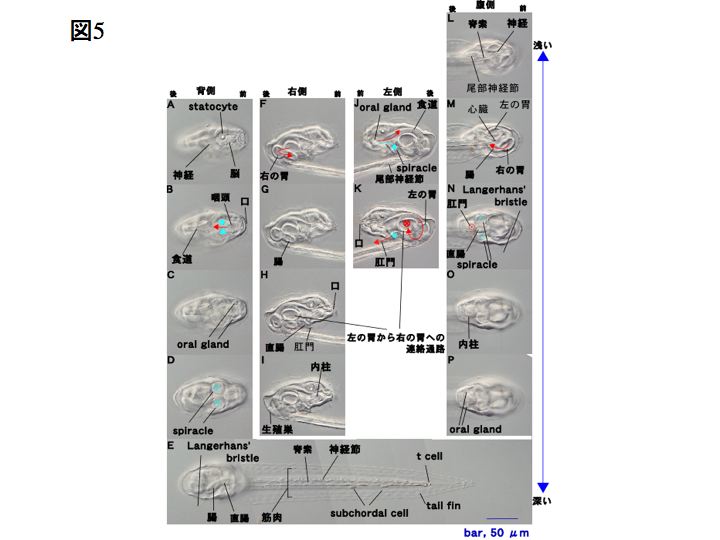
ワカレオタマボヤの幼若体の構造
下図は変態直後の幼若体(Stage6)の構造です。顕微鏡の焦点面をずらして撮影してあります。
Stage6の幼若体は生殖巣が未発達ですが、それ以外では成体と同じ構造を有しています。
オタマボヤの尾部の構造は、同じ尾索動物亜門のホヤ類の幼生と比較すると体幹部に対して90度ずれています。このためホヤ類の幼生とは泳ぎ方が異なります。このずれは、オタマボヤの初期卵割過程で形成されると考えられます。
オタマボヤのハウス
オタマボヤの成体はハウスと呼ばれる構造物を表皮から分泌します。ハウスは餌となる植物プランクトンを収集する機能を持っています。餌の収集には尾が重要な働きをします。オタマボヤは尾を前後に振り、ハウスへの水流を作り出します。水流により胴部後方の左右に位置する入水口からハウスの中へと餌などの微粒子を含んだ海水が流入します。ハウス内のフィルターを通過した海水は出水口から流出し、一方でフィルターにトラップされた微粒子はオタマボヤの口へと運ばれます。ワカレオタマボヤの1個体は毎時最大12.5mlの海水を濾過することが分かっています(Alldredge 1981)。
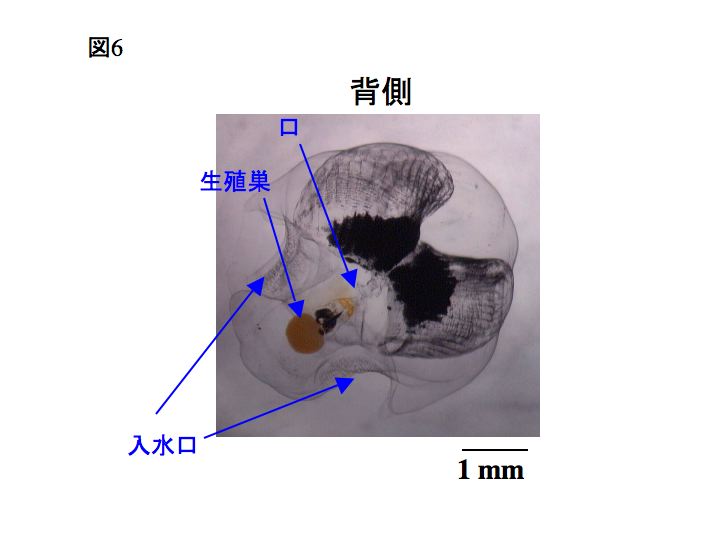
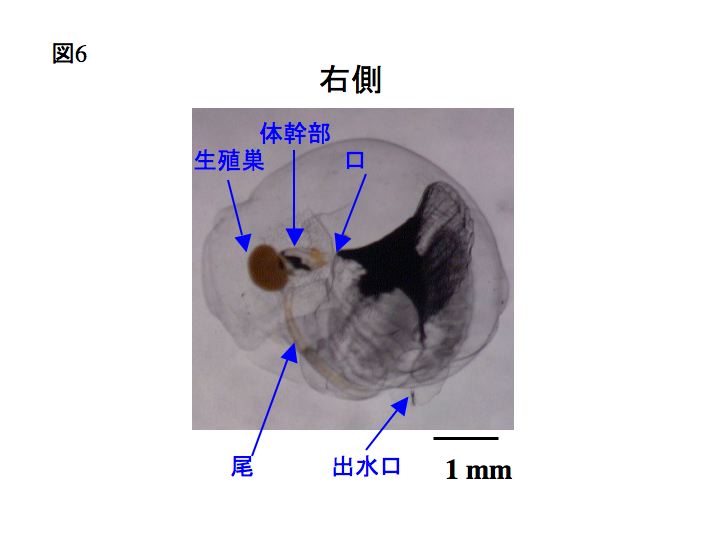
下の写真では、ハウスが分かりやすくなるよう、オタマボヤを黒色のインクを薄めた海水の中に入れてあります。黒インクが濃縮された部分がハウスの餌収集フィルター部分で、成体の口へとつながっています。茶色く楕円に見えるのは生殖巣です。スケールバーは1mm。
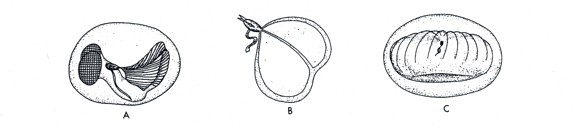
オタマボヤのハウスの形は種によって様々です。
下図では、A: オタマボヤ科、B:サイヅチボヤ科、C:カサオタマボヤ科のハウスの形を示しています。
(動物系統分類学8(下)中山書店より)
オタマボヤはハウスを1日に数回作り替えます。新しいハウスは、古いハウスの中にいる時にすでにハウス形成組織上に形成されています。オタマボヤは古いハウスを脱ぎ捨てると、数十秒から数分泳いだのち新しいハウスを広げます。ハウス形成時には、段階に応じて泳ぎかたが変化します。ハウスの作りかえは数分で完了します。
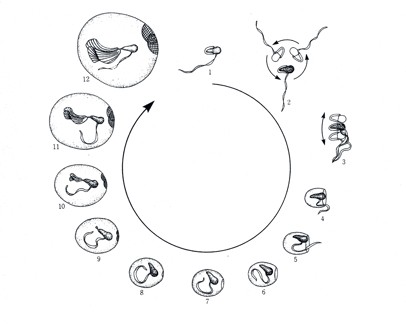
(動物系統分類学8(下)中山書店より)
1-5 ワカレオタマボヤの発生について
ワカレオタマボヤの成体は成熟するとハウスから抜け出し、海水中で放卵または放精を行います。地中海では明け方に産卵することが報告されています。雌も雄も放卵・放精後、すぐに死んでしまいます。オタマボヤの卵の直径は約75-107μmで、卵は1個体の雌から5〜300個できます。精子は全長約30μmで、頭部は3μm、幅1μmです。精子は卵に向かって泳ぎます。
ワカレオタマボヤは卵や胚が透明なため、顕微鏡での観察に適しています。私たちは、ワカレオタマボヤが卵から成熟した成体になるまでをノマルスキー顕微鏡で観察しました。以下では、受精から孵化まで・孵化から変態まで・変態から生殖巣成熟までの3つの段階に分けて発生過程を説明します。
受精から孵化までの発生過程
ワカレオタマボヤの初期発生は非常に早く、13℃で育てると受精後6時間でオタマジャクシ型幼生が孵化します(3-1参照)。原腸陥入までの観察は「初期卵割過程の観察」で詳しく説明しています。原腸陥入は32細胞から始まり64細胞期に完了します。原腸陥入したのち、3.5時間目には頭部と尾部の間にくぼみができ始め、頭尾の区別ができるようになります。
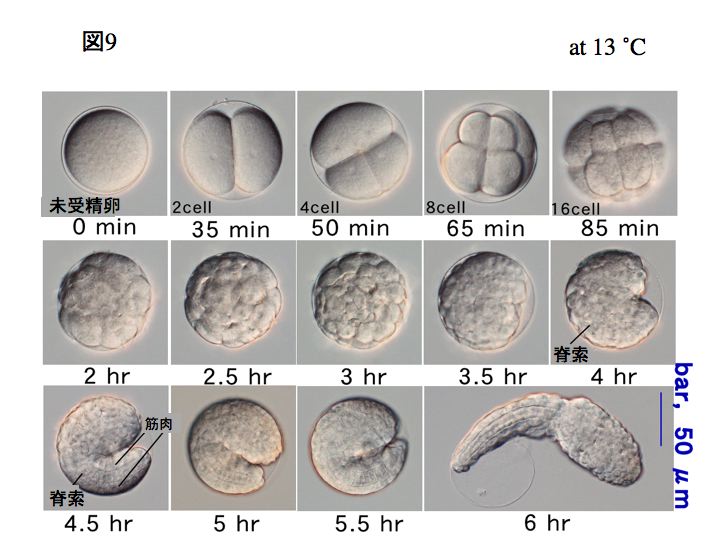
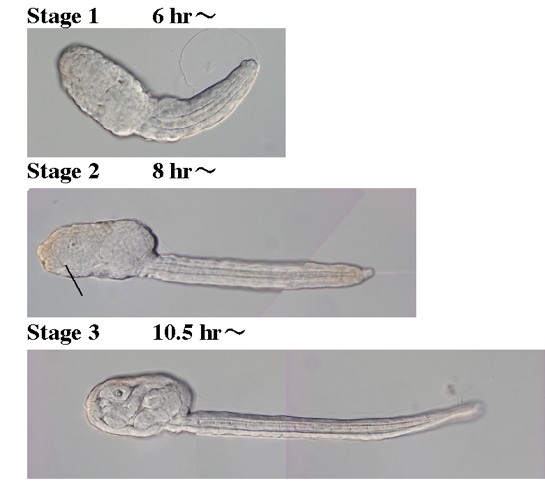
孵化から変態までの発生過程
孵化後の幼生の発生段階はStage1からStage6に分けられています。
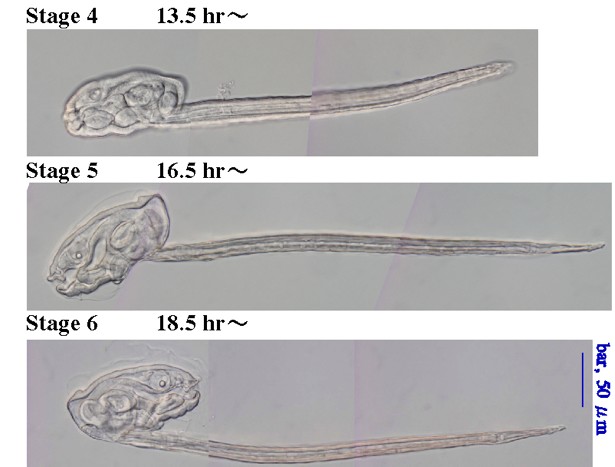
変態後の発生過程とステージ
変態が完了すると、幼若体はハウスを作るようになります。変態後の幼若体は、生殖巣が未熟である以外ほぼ成体と等しい構造をしています。変態後、オタマボヤの体細胞は細胞分裂せずDNA複製だけを行い、polyploid(倍数体)になります。最終的には1000倍体程度までDNAが増加します(Ganot,2002)。
1-6 ワカレオタマボヤのゲノムプロジェクトについて
ワカレオタマボヤのゲノムDNAの配列は、ノルウェーのSars研究所とEMBLの共同でほぼ明らかにされました。ワカレオタマボヤのゲノムサイズは64Mb(哺乳類の1/50,ホヤの2/5)で、脊索動物で調べられたゲノムサイズの中ではもっとも短いことが分かっています。遺伝子はゲノムの5kbごとに約1つの密度で分布しており、遺伝子数は約15,000と見積もられています(Seo et al.,2002)。
1-7 役に立ちそうなページ
- European Appendicularians
- Sars International Centre for Marine Molecular Biology
- Genoscope
- Marine Observatory at Villefranche-sur-Mer (CNRS)
- 京都大学フィールド科学教育センター 瀬戸臨海実験所
参考文献
- Hirst A.G. et al. (2003) Adv. Mar. Biol. 44, 1-142.
- Shelbourne J.E.(1962) J. Mar. Biol. Ass. U. K. 42, 243-252.
- Alldredge A.L. (1972) Science 177, 885-887.
- Seki H. (1973) La Mer. 11, 153-158.
- Alldredge A.L. (1982) Bull. Mar. Sci. 32, 250-254.
- Nishino A. and Satoh N. (2001) Genesis 29, 36-45.
- Alldredge A.L. (1981) Limnol. Oceanogr. 26, 247-257.
- Ganot P. and Thompson E.M. (2002) Dev. Biol. 252, 59-71.
- Seo H.C. et al. (2002) Science 294, 2506.
- Bone Q. (1998) The Biology of Pelagic Tunicates, New York: Oxford University Press.
- Galt C.P. and Fenaux R. (1990) Urochordata-larvacea, In Reproductive Biology of Invertebrates (ed. Adiyodi K.G and Adiyodi R.G.), p471-469. New York: John Wiley & Sons.
- 西尾孝也(2003)修士論文 オタマボヤの飼育方法の確立と初期発生の記載
- 岩波生物学辞典 第四版 岩波書店
- 動物系統分類学8(下)半索動物・原索動物(1986)p318-337 中山書店
- 日本海洋プランクトン図鑑 保育社