3-3 トランスポゾンを用いた突然変異体の作製とその技術の開発
全ての生命現象は、遺伝情報の最小単位である遺伝子から生じます。突然変異(単に変異とも言います)とは、この遺伝子を構成する遺伝物質であるDNAやRNAの塩基配列の変化(置換および挿入・欠失)を意味します。全ての生物において突然変異は普遍的に起こるものであり、それは細胞分裂時のDNA複製で生じるエラーや、化学物質・放射線照射、ウイルスの感染、トランスポゾンの転移などによって偶発的に、減数分裂時などには意図的に引き起こされます。多くの突然変異は遺伝子の機能に影響を及ぼしませんが、遺伝子内の重要なアミノ酸配列や遺伝子の転写量を変化させるような突然変異が生じると、その遺伝物質を持つ細胞の形質は変化します。また、体細胞に生じた突然変異は子孫には遺伝せず、生殖細胞に生じた突然変異のみが子孫に代々伝わることになります。そのため「体細胞に突然変異は生じていないが生殖細胞に“効果的な”突然変異を持つ」個体がいたとすると、その個体を親とする子孫は親とは異なる形質をもち、それは表現型として現れます。これが一般的に突然変異体と呼ばれるものです。
1926年にH. J. MullerがショウジョウバエにX線を照射して突然変異を誘発できることを実験的に証明して以来、生物学では人為的な突然変異の誘発によって得られた突然変異体の表現型を観察することで、様々な生命現象に関わる重要な遺伝子やメカニズムをつきとめようとしてきました。多くの生物のゲノムの解析が進んだ今では、この遺伝学的手法は生物学において欠かせないものとなっています。特に、ショウジョウバエや酵母菌、線虫、ゼブラフィッシュ、マウス、シロイヌナズナ、ファージといった生物は、生物学的・経済的に扱いやすくモデル生物と呼ばれ、生物学(遺伝学)の分野において普遍的な生命現象の研究に盛んに用いられています。
当研究室で扱っているワカレオタマボヤは、継代飼育できること、ライフサイクルが短いこと、ゲノムがコンパクトで遺伝子間距離が短いこと、遺伝子重複があまりないことなどから、突然変異体の作製と解析に適した実験動物であると考えられます。オタマボヤと同じ脊索動物亜門に属するカタユウレイボヤ(Ciona intestinalis)では既に、トランスポゾン導入による突然変異体の作製と解析が進んでいます(Sasakura et al., 2005)。トランスポゾンとは、ゲノム上の位置を転移することができる塩基配列のことで、その一部にはトランスポゾン独特の配列をもちます。このため、トランスポゾンを用いると、ゲノム挿入後の再転移によって更なる変異を誘発することや、変異した遺伝子を容易に特定することが出来ます。ホヤで行われているこの変異体作製法は、近縁種であるオタマボヤにもおそらく適用できると考えられ、当研究室ではSasakuraらの方法を元にして突然変異体の作製を始めています。
*トランスポゾンについて
トランスポゾンは、動く遺伝子や転移因子とも呼ばれる、DNA上の位置を転移(トランスポジション)できる塩基配列のことを言います。トランスポゾンが入り込んだ場所の遺伝子が、トランスポゾンの挿入により壊れてしまうことがあるので、これを突然変異体を人為的に作製する手段として利用することができます。トランスポゾンにはDNA配列が直接転移するもの(狭義のトランスポゾン)と、転写されたRNAが逆転写を経て転移するもの(レトロトランスポゾン)の2種類がありますが、ここでは前者についてのみ解説します。
トランスポゾンが転移するためには、転移酵素と呼ばれる酵素が必要です。普通、トランスポゾンは自身の塩基配列中にこの転移酵素の遺伝子配列を持っており、細胞内の様々なタンパク質の力を借りて転移酵素は転写・翻訳されます。一方、トランスポゾンの塩基配列の両端には、トランスポゾン独特の配列である逆向き反復配列が存在します(図1の赤矢尻)。転移酵素はこの逆向き反復配列を認識して、トランスポゾンをDNAから切り取ります。このとき、元々トランスポゾンが入っていた場所には、footprintと呼ばれる逆向き反復配列一組(もしくはその一部)が残ります。切り取られたトランスポゾンと転移酵素の複合体は他のDNA上へ移動し、転移酵素はトランスポゾンをDNAへ挿入します。(図1)
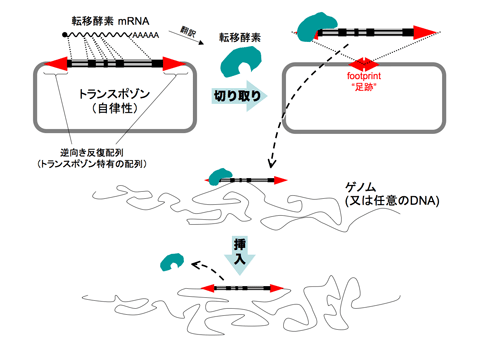
図1 トランスポゾンがはたらく仕組み
このように、自身の存在だけで完全な活性を持つトランスポゾンの状態を「自律性である」と言います(図2上段)。ここで、例えば、トランスポゾンが持っている逆向き反復配列はそのままで転移酵素の遺伝子配列の一部を削ってしまうと、転移酵素は翻訳されなくなるか正常な転移酵素が作られなくなります。すると、トランスポゾンは転移することが不可能になってしまいます(図2下段右上)。しかし、そこへ翻訳されれば正常な転移酵素ができるmRNAを加えてやると、このmRNAからつくられた正常な転移酵素がトランスポゾンの逆向き反復配列を認識するため、トランスポゾンは転移可能になります(図2下段右下)。このように、自身の存在だけでは活性が無く、活性化には他からの助けを必要とするトランスポゾンの状態を「非自律性である」と言います。
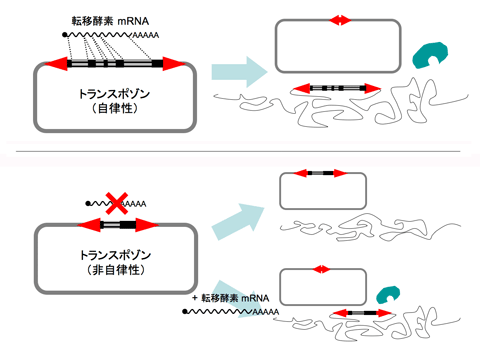
図2 自律性(上段)・非自律性(下段)トランスポゾン
このようなトランスポゾンの仕組みを応用して、多くの研究者が突然変異体を作製しています。具体的には以下のような方法をとります。
目的の動物の突然変異体を解析するには、突然変異が一カ所にのみ起こっていること、またその突然変異が以降変化しないことが重要です。そのためには、トランスポゾンが自律的に動くことでDNAが次々に変化していくのを防ぐため、逆向き反復配列を残した非自律性のトランスポゾンを使います。非自律性トランスポゾンの塩基配列の中には、トランスポゾンが動物のゲノムDNAの中に挿入されたかどうか確認できるように、モニター用の蛍光タンパク遺伝子(例:GFP、venus-YFP)などの任意の遺伝子を組み込みます。(図3上段)
この蛍光タンパク遺伝子を持つ非自律性トランスポゾンを、正常な転移酵素のmRNAと一緒に動物の卵細胞に入れて、通常の精子と受精させます。この受精卵が幼生、成体へと成熟すると、多くは通常通りの個体となりますが、中には体の一部が蛍光で光る個体が現れます。この蛍光が、トランスポゾンがDNAに組み込まれた可能性があるという印になります。この光る個体の生殖細胞を通常の個体の生殖細胞と受精させ、生まれてきた次世代も蛍光で光っていれば、代々生殖細胞系列に蛍光タンパク遺伝子を持つトランスポゾン配列が組み込まれたトランスジェニック(遺伝子組み換え)個体が得られたことになります。(図3下段)
このトランスジェニック個体の大半は単に正常な個体が光るものですが、稀に表現型が通常と異なる(例えば伸びるはずの尾が伸びないなど)系統が現れます。この表現型異常が突然変異体です。こうして得られた突然変異体のゲノムDNAを調べるときに、図1で示した逆向き反復配列や自分が組み込んだ任意の遺伝子配列を手がかりに、トランスポゾンが挿入された遺伝子、つまり変異の生じた遺伝子を特定することができます。
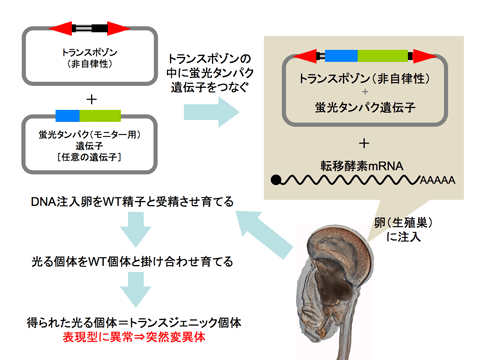
図3 トランスポゾンを用いて突然変異体を得る方法
トランスポゾンを用いた突然変異体作製法は、蛍光タンパクだけでなく任意の遺伝子を自由に組み込むことができます。また、一度ゲノムDNAにトランスポゾンが組み込まれれば転移酵素のmRNAを注入するだけで再び転移を生じさせ突然変異を誘起することが可能です。さらに上述のように、変異した遺伝子の特定も容易です。当研究室はこの方法を用いてワカレオタマボヤのトランスジェニックライン・突然変異体作製に取り組んでいます。
参考文献
- Muller H. J. (1928) Proc Natl Acad Sci U S A. 14(9), 714-726.
- Sasakura Y. et al. (2005) Proc Natl Acad Sci U S A. 102, 15134-15139.